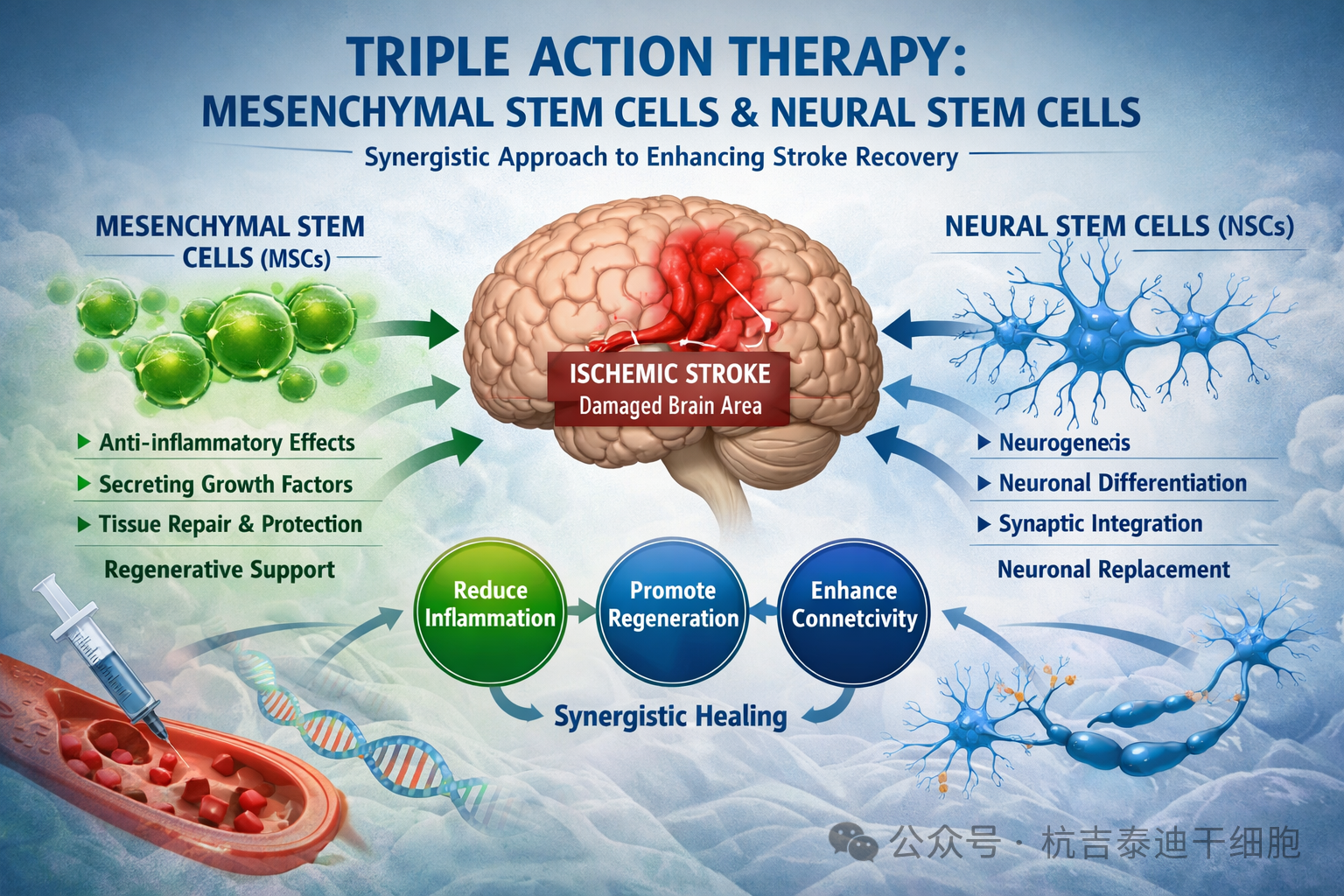
脑神经干细胞(NSCs)被定义为早期神经上皮组织中具有自我更新能力且能分化为多类神经元后代的细胞,可生成神经元、星形胶质细胞和少突胶质细胞。这类细胞主要存在于发育中的大脑中,在成年大脑的脑室下区和海马区也有少量分布,可在体外进行分离与扩增。
一、神经科学中的神经干细胞简介
神经干细胞(Neural Stem Cells, NSCs)具有两个核心特征:无限的自我更新能力和多向分化潜能,即能够分化为中枢神经系统(CNS)的三种主要细胞类型——神经元、星形胶质细胞和少突胶质细胞。神经干细胞是神经系统中分化程度最低的细胞,由于缺乏足够特异的免疫细胞化学标记,因此主要通过功能特征来定义。
20世纪80年代末至90年代初,科学家在啮齿类动物脑内发现了成体神经干细胞,随后又在成人大脑中得到证实,这一发现推翻了“神经发生在发育完成后终止”的长期教条。神经干细胞在整个生命过程中持续存在于中枢神经系统中,通过不断产生新的神经元和神经胶质细胞,参与脑的发育与维持。
在发育中及成体哺乳动物的大脑中,الخلية الجذعية العصبية位于称为神经发生微环境(neurogenic niche)的特殊区域中,这些微环境调控着干细胞的增殖、分化和维持。两个主要的神经发生区包括:
-
侧脑室下区(SVZ):位于侧脑室壁;
-
海马齿状回下颗粒区(SGZ):位于海马体。
此外,研究还发现下丘脑等区域也存在潜在的神经发生“生态位”。这些区域为神经干细胞提供了有利的微环境,支持其自我更新及向神经元、星形胶质细胞乃至少突胶质细胞的分化。神经干细胞在神经发生中发挥核心作用,其产生新神经细胞的能力正是其在再生医学及神经疾病治疗中具有重要意义的基础。

二、脑神经干细胞的生物学特征与调控机制
神经干细胞起源于神经板中的神经上皮细胞(neuroepithelial cells),这些细胞通过对称分裂扩增后,逐渐转化为放射状胶质细胞(radial glia)。放射状胶质细胞是真正的神经干细胞,在体内可生成神经元和神经胶质细胞。
神经干细胞的两个核心属性是无限自我更新和多向分化潜能,能够产生包括神经元、星形胶质细胞和少突胶质细胞在内的全部主要神经谱系。与其后代——**祖细胞(progenitors)**的区别在于:祖细胞的分化潜能更受限,而神经干细胞具备较强的可塑性。
在发育过程中,神经干细胞经历对称与非对称分裂:
-
对称分裂可扩大干细胞库;
-
非对称分裂则产生一个干细胞与一个分化的祖细胞,进而促进神经元多样性。
在成体脑中,神经干细胞主要位于侧脑室下区(SVZ)与海马齿状回下颗粒区(SGZ),并能在整个成年期持续存在。
识别神经干细胞常用的分子标志包括:
- 内斯廷
- Sox2
- 武藏1
但这些标志并非神经干细胞所独有,也可在祖细胞或成熟神经细胞中表达。
神经干细胞及其后代呈现出明显的异质性,不同区域的细胞群具有独特的分子特征和功能标记。
调控神经干细胞维持、增殖及命运决定的关键信号通路包括:
-
Wnt/β-catenin
-
缺口
-
索尼克刺猬(嘘)
-
骨形态发生蛋白(BMP)
转录因子Sox2与TLX对自我更新和分化至关重要:
-
Sox2促进干细胞增殖与神经元成熟;
-
TLX维持其未分化状态。BMP信号促进干细胞静息及星形胶质分化,而其拮抗物Noggin可促进干细胞自我更新与神经发生。
细胞外基质(ECM)与血管微环境同样通过结构支持与信号分子传递调控神经干细胞行为,其中的层粘连蛋白(laminins)、tenascins及蛋白多糖(proteoglycans)参与细胞黏附、迁移与生长因子信号调节。
在代谢与表观遗传层面,神经干细胞倾向于使用糖酵解代谢;其静息与激活状态受到营养与代谢信号调控。表观遗传机制(如染色质重塑与非编码RNA)在神经干细胞可塑性与谱系分化中也起关键作用。神经干细胞具备惊人的可塑性,能够动态响应内外部信号的变化。

三、神经干细胞在神经发生、脑发育与可塑性中的作用
神经干细胞是神经系统的“原始种子细胞”,在发育过程中负责为大脑补充神经元与神经胶质细胞。发育完成后,其数量显著下降,但仍有部分细胞在成体期持续生成新的神经元与胶质细胞。
在胚胎发育阶段,神经干细胞最初以神经上皮细胞形式存在,通过对称分裂扩增前体细胞库,随后转化为放射状胶质细胞,通过非对称分裂产生神经元及神经前体,随后形成胶质细胞及其前体。
这一过程最终构建出整个中枢神经系统(CNS)的神经元与神经胶质网络。
在成年脑中,神经干细胞主要位于:
-
侧脑室下区(SVZ):生成的神经母细胞沿嗅球迁移通路(RMS)迁移至嗅球,分化为中间神经元;
-
海马齿状回下颗粒区(SGZ):生成的前体细胞在局部迁移并分化为齿状回颗粒神经元。
这些新生神经元可整合入原有神经环路,参与海马相关的学习与记忆过程。
成体神经发生受多种内外因素调控,包括生态位信号、生长因子及细胞间相互作用。随着年龄增长,神经干细胞的增殖、分化及新生细胞的存活率逐渐下降,而干细胞静息状态增加是关键原因。
运动、环境丰富化等生理刺激可促进神经发生,而应激及病理状态(如癫痫)则会调节这一过程——癫痫发作在急性期可短暂促进神经发生,但长期则导致神经再生受损。
神经发生(Neurogenesis)使神经系统获得新的神经元来源,增强神经可塑性。神经发生微环境由信号分子、细胞外基质及细胞间相互作用组成,决定着干细胞的自我更新、分化与神经发生能力。
四、神经干细胞在神经系统疾病与损伤中的作用
在成人脑中,神经干细胞主要存在于侧脑室下区(SVZ)与海马齿状回下颗粒区(SGZ),可自我更新并分化为神经元、星形胶质细胞与少突胶质细胞。
在神经退行性疾病中,如阿尔茨海默病(AD)、帕金森病(PD)与多发性硬化(MS),神经干细胞的增殖与神经发生均受损:
-
AD患者的前体细胞增殖能力下降;
-
PD中多巴胺能神经元缺失导致前体细胞数量减少。
在中风(卒中)及创伤性脑损伤(TBI)等中枢神经损伤后,内源性神经干细胞被激活并迁移至损伤区,但新生神经元的存活率极低,例如在缺血性损伤大鼠模型中,仅约0.2%的受损神经元被替代。
损伤区形成的胶质瘢痕会限制神经干细胞的分化与整合,使其更易分化为胶质细胞而非神经元。神经干细胞功能障碍(如自我更新能力下降、分化受阻)会加重病理状态,并受老化、炎症及表观遗传异常影响。在癫痫等疾病中出现的异常神经发生可能导致新生神经元迁移与整合失败,从而增加癫痫复发风险。
神经干细胞还能与中枢神经系统免疫反应互作,调节炎症并促进修复。研究表明,移植的神经干细胞可:
-
抑制炎症反应;
-
减少免疫细胞浸润;
-
分泌促血管生成及修复的营养因子。
其修复机制涉及Notch、Wnt/β-Catenin及脑源性神经营养因子(BDNF)等信号通路。然而,干细胞疗法的疗效仍受限于细胞存活率低、分化受限及抑制性微环境等因素。
五、神经干细胞的治疗应用、挑战与未来方向
神经干细胞作为多能前体细胞,具有自我更新及分化为神经元、星形胶质细胞与少突胶质细胞的能力,并在一生中持续存在于如SVZ与齿状回等神经发生区中。
这些细胞可来源于:
-
胎儿组织
-
胚胎干细胞(ESCs)
-
诱导多能干细胞(iPSCs)
-
成人来源细胞
不同来源在移植治疗中的优势与挑战各异。
神经干细胞适合进行基因工程改造,常用技术包括病毒载体转染、脂质体转染、电穿孔及磷酸钙共沉淀等,可用于体外修饰以表达治疗性分子。
CRISPR/Cas9基因编辑技术已广泛应用于神经干细胞与iPSCs,可实现精确基因校正或疾病突变修复。
干细胞治疗的主要挑战包括:
-
免疫排斥反应
-
肿瘤形成风险
-
伦理争议
移植可能引发宿主免疫反应导致移植物破坏,解决策略包括使用患者特异性干细胞及免疫抑制疗法。多能干细胞来源的神经干细胞可能具有肿瘤形成风险,因此需严格安全评估。胚胎或胎儿来源细胞引发伦理问题,而iPSC来源细胞可在一定程度上避免。
另一个关键挑战是确保移植细胞的正确分化与整合——损伤环境往往不利于神经元或少突胶质分化,未分化细胞还可能带来肿瘤风险。
调控内源性神经干细胞促进修复的策略包括:
-
外源性施用生长因子,如VEGF、EGF、FGF2,可刺激体内干细胞增殖与分化。
当前研究趋势是通过联合疗法整合神经干细胞移植与生物材料、基因治疗及康复训练,以提升细胞存活率、分化效率与功能整合。生物工程方法(如三维支架与生物材料)正用于优化神经微环境,引导干细胞生长与分化。
写在最后:站在再生医学的新起点,神经干细胞疗法的目标已不仅仅是证明概念,而是如何将其转化为稳定、可及的临床解决方案。未来的核心任务在于优化细胞制备的标准化流程、攻克移植后的整合效率难题,并通过联合治疗策略最大化功能恢复。随着临床证据的不断累积与治疗方案的规范化,我们有理由期待,神经干细胞疗法将逐步从前沿研究走向临床实践,重塑神经系统疾病的治疗格局,为患者带来切实的治愈曙光。
郑重声明:本文版权归原作者所有,转载文章仅为传播更多信息之目的,如作者信息标记有误,请第一时间联系我们修改或删除,多谢。
اترك تعليقاً